
Viz také:
V předchozím díle minisérie jsem ukázal, jak se zrodila hypotéza o "běžcích" coby polyfyletickém seskupení ptáků v rámci monofyletické skupiny Palaeognathae, a vyjmenoval jsem nedlouhou řádku studií, které tento závěr zatím podpořily. Zatím jsem ale neukázal důvod, proč by tento scénář měl být důvěryhodnější než všechny alternativy, zvláště když morfologické i molekulární studie pro "běžčí" monofylii v předchozích dekádách snesly tolik důkazů. To by měl článek níže napravit.

1. Smith et al. 2012: fylogenetika, metody a problémy
Skupina autorů z týmu Early Bird se rozhodla ověřit otázku asi ze všech nejdůležitějších, totiž jak dobře si vlastně hypotéza "běžčí" polyfylie stojí. Důvodů k pochybám je několik. Zaprvé existuje několik studií jaderných genů, které monofylii skupiny "Ratitae" podpořily (např. již citovaní García-Moreno & Mindell 2000). Zadruhé jednotlivé "běžce" stále spojuje velké množství odvozených morfologických znaků, především pak z postkrania, a ačkoli je lákavé odvysvětlit je neotenií, nikdy nebylo prokázáno, že neotenie dokáže v případě "běžců" vyprodukovat dostatečně mnoho konvergencí k tomu, aby zkreslovala výsledky fylogenetických analýz. Zatřetí ani datasety tak velké, jako jsou ty sestavené týmem Early Bird (Hackett et al. 2008; Harshman et al. 2008), nejsou zárukou dosažení správných výsledků. Např. postavení žebernatek v rámci metazoí je případem, kdy i velmi velké datasety podávají rozporuplné výsledky. Zvětšování datasetu je dobrou strategií pro eliminaci náhodného šumu, ale v molekulární evoluci je zdokumentováno mnoho jevů, které mohou výsledky analýzy zkreslit nenáhodně, tj. určitým směrem. Dvěma takovými biasy jsou přitahování dlouhých větví (long branch attraction) a konvergence na podobném zastoupení bází v sekvenci (base composition bias) a právě pomocí nich zastánci "běžčí" polyfylie vysvětlují, proč monofyletičtí "běžci" vycházeli i z tolika molekulárních studií: tinamy jsou dlouhá větev a přitahují se k outgroup, pštrosi zase sdílejí nozký obsah tyminu s cytozinem v mitochondriálním genomu s kivii a kasuárovitými.
Krom těchto jevů ale může fylogenezi zkreslit i nesoulad mezi genovými stromy (gene trees) a druhovými stromy (species trees) v důsledku nekompletního třídění linií. Smith et al. (2012) je sice necitují, ale Baker & Haddrath (2010) v abstraktu z předloňského ornitologického kongresu navrhli, že by "běžčí" polyfylie mohla být artefaktem právě tohoto jevu. Na tomtéž místě zmiňují, že když na rozsáhlý, dosud nepublikovaný dataset aplikovali pokročilé bayesovské metody pro rekonstrukci druhového stromu místo prostého zřetězení (concatenation) sekvencí jednotlivých genů, vyšli jim "běžci" zase monofyletičtí. A s tím, jak poznáváme lépe a lépe zákonitosti genomové evoluce, nacházíme nové a nové problémy. Co třeba heterotachie, tedy jev, při němž se rychlost substituce na jediné nukleotidové pozici mění v čase (a tedy napříč fylogenetickým stromem)? Nechceme-li se vzdát výhod parametrických metod, musíme využít modelů, které věrohodnost pro každý znak počítají jako vážený součet přes několik množin délek větví (mixture models). Jenže Matsen & Steel (2007) ukázali, že analýza dvou genů, jejichž fylogenetické stromy se liší pouze v délkách větví (tj. topologie genových stromů je stejná), nám pod "mixture model" může vyhodit strom s topologií úplně odlišnou. Problémy jsou i s alignmentem: fylogenetická analýza špatně zarovnaných sekvencí vyhodí špatný strom. A nemělo by se zapomínat ani na to, že podpora jednotlivých uzlů je skoro důležitější než samotný bodový odhad stromu, takže teoretičtí fylogenetici se vyžívají v hledání situací, kdy naprosto chybné klady obdrží vysoké hodnoty bootstrapu a posteriorních pravděpodobností. Co když je drasticky nadhodnocena např. vysoká podpora pro nepštrosí paleognáty v analýzách Harshmana a spol.?
Smith a spol. proto otestovali hypotézu "běžčí" polyfylie jednou z nejjednodušších metod vůbec: vzali nezávislá data (tj. ta, která nebyla zahrnuta v žádné z analýz, která původní hypotézu podpořila) a podívali se, co z nich vyleze. Podobné testování kontroverzních hypotéz má v ptačí systematice tradici: Morgan-Richards et al. (2008) se takto podívali na zoubek kladu Metaves skrze mitochondriální data (a hypotézu zamítli), Wang et al. (2011) loni zase s 30 novými jadernými geny ověřovali existenci skupiny Eufalconimorphae, složené z pěvců, papoušků a sokolů (hypotézu v plném rozsahu potvrdili, ale ukázali, jak zoufalá je situace v případě myšáků). Smith a spol. jsou ještě o něco ambicióznější: předkládají hned 40 zbrusu nových lokusů roztroušených po celém jaderném genomu. Dohromady dávají přes 22 000 bází, což konkuruje původnímu datasetu Harshmana a spol., a lze mezi nimi najít i "anonymní lokusy", tzn. náhodné fragmenty genomu, které mohou zahrnovat i dlouhé introny a mezigenové regiony. Pokud by tyto oblasti genomu nesly jiný signál než exony, krátké introny a UTRs použité Harshmanem a spol., Smith et al. (2012) by to dokázali zjistit.
Zarovnávání sekvencí Smith a spol. provedli manuálně ("eyeball alignment"), přičemž problematické sekvence bez milosti vyřazovali. Zároveň ale vyzkoušeli i trochu jiný postup: aby prověřili, jak moc je analýza na zarovnání citlivá, nakrmili program PRANK (provádějící alignment automaticky na základě maximální věrohodnosti) 15 tzv. "guide trees". Jelikož se PRANK snaží určit, za kterou mezeru (gap) v zarovnané sekvenci je zodpovědná delece a za jakou inzerce, potřebuje strom – "guide tree" –, na němž by mohl zrekonstruovat ancestrální sekvence. Když se mu žádný nezadá, PRANK si ho sestaví sám skrze neighbor-joining, ale Smith et al. (2012) této možnosti nevyužili. Místo toho mu postupně zadali všechny stromy, které uznali za realistické: "realističnost" přitom předpokládá monofylii nanduů (Rhea + Pterocnemia), monofylii tinam (Tinamus + Crypturellus) a pevně fixovanou topologii outgroup (která předpokládá monofylii hrabavých a drůbeže, čili opět nic kontroverzního). Proměnlivé byly jen vztahy pštrosů, nanduů, tinam a emu, což dá výše zmíněných 15 stromů. Délky jejich větví autoři spočítali pomocí věrohodnostní analýzy nad manuálním alignmentem.
Pokud jde o hlavní analýzu, byli Smith a spol. velice opatrní. Konstatují, že obrovské mnohalokusové datasety sice slibují mnohé, ale jen tehdy, pokud nespadají do problematické oblasti parametrového prostoru: narážejí na "anomální zónu", kterou popsali Degnan & Rosenberg (2006) a která má potenciál být fylogenetiky vyslovována se stejným nádechem zlověstnosti, jenž už léta doprovází zónu Felsensteinovu. V této zóně mají druhové stromy takovou kombinaci krátkých vnitřních větví, která vygeneruje genové stromy odporující stromu druhovému častěji než ty, které se s druhovým stromem shodují. Kubatko & Degnan (2007) pak ukázali, že k tomu, aby se maximální věrohodnost stala při rekonstrukci druhového stromu ze zřetězených sekvencí statisticky nekonzistentní, se stačí k obávané zóně jen přiblížit. Smith s kolektivem tedy proto kromě zřetězeného datasetu analyzovali metodami nejvyšší parsimonie a věrohodnosti i každý lokus zvlášť, přičemž věrohodnostní analýzu ještě provedli jak s jedním modelem naraženým na celý dataset, tak i se samostatným modelem pro každý lokus. Trojku hlavních fylogenetických metod doplnila bayesovská inference, pod níž lze navíc nasadit modely adresující výše zmíněný problém heterotachie. Smith et al. (2012) vyzkoušeli hned dva: jednak "mixture model" popsaný výše, jednak "covarion model", který se heterotachii pokouší obelstít trochu jinak: používá skrytý Markovův proces*, který dovoluje, aby se nukleotidové pozice během své evoluce přepínaly ze stavu "off" (kdy na nich žádné substituce neprobíhají) do stavu "on", kdy k mutacím dochází normálně (Kolaczkowski & Thornton 2008). Tato celkem jednoduchá původní myšlenka Tuffleyho a Steela (1998) byla časem doplněna řadou rafinovaných vymyšleností, takže původní binární volba "on/off" byla rozšířena na výběr z vícero rychlosntních kategorií (takový model vlastně dělá s variací v tempu substituce napříč časem totéž, co klasický gama parametr s variací v tempu substituce napříč nukleotidovými pozicemi) a posléze i kombinaci obou předchozích možností.
*To, že je Markovův, znamená, že pravděpodobnost další události nezávisí na událostech předchozích. V jazyce molekulární evoluce to znamená, že důležité je jen to, jaký je stav nukleotidové pozice teď, ne to, jaký byl předtím. To, že je skrytý, znamená, že jeho jednotlivé stavy nemůžeme přímo pozorovat – tyto "skryté" stavy jsou ale nějakým mechanizmem propojeny s pozorovatelnými výstupy a lze je z nich odhadnout.
Rozpory mezi genovými a druhovými stromy Smith a spol. zjišťovali pomocí programu BUCKy, který bayesovsky (dvojnásobnou aplikací Markovových řetězců Monte Carlo) odhaduje tzv. faktory konkordance (shody), tedy procento genomu, které podporuje zadaný klad. Jako každá bayesovská metoda, i BUCKy vyžaduje specifikování priorní pravděpodobnosti: zde jde o parametr α, který vyjadřuje apriorní přesvědčení systematika o tom, jak moc si budou jednotlivé genové stromy z jeho datasetu navzájem odporovat (α = 0: vůbec, α = ∞: historie jednotlivých genů budou absolutně nezávislé). S priorními pravděpodobnostmi je vždycky trochu potíž: ve fylogenetice třeba panuje shoda na tom, že všechny topologie by měly mít stejný prior – žádný strom bychom neměli upřednostňovat dřív, než analyzujeme data. Podstatně kontroverznější je volba priorů pro délky větví, a bayesovsky založení biologové si stěžují, že největší výzkum v oblasti priorních pravděpodobností se soustředí na to, jak je učinit neinformativními (Ronquist & Deans 2010). Ze Smithe a jeho spoluautorů by ale Ronquist s Deansem měli radost: jejich alfa je informativní, odvozená z předchozích studií paleognátí fylogeneze (Harshman et al. 2008; Phillips et al. 2010) a poskytující následující roztomilé priorní rozdělení pro počet odlišných genových stromů:


Autoři použili BUCKy ke dvěma účelům: jednak aby určili, kolik lokusů podporuje ten který klad, jednak k rekonstrukci druhového stromu pomocí "kvartetové metody", která má tu příjemnou vlastnost, že je statisticky konzistentní a lze garantovat, že správnou topologii druhového stromu najde i tehdy, když neodpovídá nejčastěji pozorovanému genovému stromu. Vyhýbá se tak výše zmíněné anomální zóně a bere v úvahu fakt, že postup, který Dawkins (2004) popsal jako "demokratické hlasování" jednotlivých genů, může být při rekonstrukci evoluční historie organizmů a populací zavádějící. Nevýhodou je pak skutečnost, že statistické konzistence může dosáhnout jen tehdy, když jsou faktory konkordance odhadnuty správně. Autoři ale dokázali obejít i tento požadavek. Rozdělili genové stromy do dvou kategorií – na ty, na nichž klad všech nepštrosích paleognátů je, a na ty, na kterých není – a pak spočítali, kolik stromů by v obou kategorií mělo být, pokud by klad nepštrosích paleognátů neexistoval (na druhovém stromě). Následně použili statistický test, aby zjistili, zda lze tuto nulovou hypotézu zamítnout.
Jako tečku za záplavou dílčích analýz Smith et al. (2012) zkombinovali svá data se sekvencemi Harshmana a spol. (2008) a Phillipse a spol. (2010), čímž složili supermatici atakující 60 kilobází. Podobný postup zvolili Wang et al. (2011) při loňském přetestování sesterského vztahu mezi pěvci a papoušky: ať už spolu jednotlivé datasety budou souhlasit nebo ne, nejrobustnějších výsledků dosáhneme, když je spojíme dohromady. Analýzy provedené nad tímto datasetem byly celkem dvě: jedna s minimálním taxon samplingem, tj. pouze s taxony společnými pro všechny dílčí datasety (pštrosa, dvě tinamy, oba nanduy a emu), druhá měla naopak taxon sampling maximální, zahrnující všechny taxony přítomné aspoň v jednom datasetu – celkem tedy pštrosa, oba nanduy, čtyři tinamy, tři ptáky moa, tři kivie, emu a kasuára. Dva kiviové a všichni moa byli v analýze reprezentováni výhradně mitochondriálními sekvencemi. Na oba tyto datasety byla nasazena parsimonie, věrohodnost i bayesovská inference; data byla opět roztříděna do několika oddílů se zvláštním modelem pro každý jeden jaderný gen. V detailech analýzy mitochondriální části datasetu Smith s kolektivem následovali to, co zvolili Phillips et al. (2010).
2. "Běžci" verzus nepštrosí paleognáti: souboj hypotéz na život a na smrt
Ve stručnosti: klad nepštrosích paleognátů (a tím pádem polyfylie "běžců") byl jednoznačně potvrzen. Nejprve autoři prozkoumali obsah 95% kredibilní množiny stromů (95% credible set of trees). Tuto množinu dostaneme tak, že ve vzorku, který sestavily Markovovy řetězce Monte Carlo (MCMC) v průběhu bayesovské analýzy, seřadíme jednotlivé stromy podle jejich posteriorní pravděpodobnosti (PP). Postupujeme od stromu s nejvyšší posteriorní pravděpodobností k méně a méně pravděpodobným stromům, dokud celková posteriorní pravděpodobnost nedosáhne hodnoty 0,95. Asi se do ní netrefíme úplně přesně (když má 1. strom PP 0,45, druhý 0,35 a třetí 0,18, celkový součet je už 0,98), ale jak ve své výborné studii o interpreaci bayesovských pojmů a měřítek ukázali Huelsenbeck & Rannala (2004), to lze napravit tím, když strom překračující 95% hranici zahrneme jen s pravděpodobností odpovídající tomu, o kolik ji překračuje. Ukázalo se, že v 95% kredibilní množině stromů se nachází jen jediná topologie, a ta obsahuje nepštrosí paleognáty. Topologie, na nichž se vyskytují monofyletičtí "běžci", byly vyloučeny dokonce z 99% kredibilní množiny stromů: ač tvoří 3 z 15 (tj. 20%) možných zakořeněných stromů popsaných výše, na posteriorní pravděpodobnosti se podílejí méně než 1%.
"Běžčí" polyfylii ve výsledcích určitě nelze svést na heterotachii, tedy změny v rychlosti evoluce za čas. Bayesovská analýza s využitím "covarion" a "mixture" modelů evoluce odhalila monofyletické nepštrosí paleognáty zase, a to s posteriorní pravděpodobností 1. Ukázalo se navíc, že MCMC pro většinu větví nezvolily víc než jednu délku (ačkoli klidně mohly). Rozbor jednotlivých genových stromů ukázal, že 24 z 38 lokusů podporuje monofylii nepštrosích paleognátů ve věrohodnostní i bayesovské analýze, zatímco pod úspornostní analýzou je to 15 z 38 (chybějící 2 lokusy nemohly být osekvenovány od jednoho z paleognátích taxonů, a tak je autoři pro teď ignorovali) – v obou případech je to víc než 16% a méně procent, které bychom očekávali, kdyby takový klad nebyl přítomen na druhovém stromě. Podle autorů může menší podpora ze strany parsimonie odrážet náchylnost metody k přitahování dlouhých větví, zde tedy konkrétně dlouhé větve tinam k dlouhé větvi outgroup. Faktor konkordance ukazuje, že monofylii nepštrosích paleognátů podporuje 93% genomu – 95% interval kredibility (tj. interval, který s pravděpodobností 95% obsahuje správnou hodnotu) je 84% až 99%. A zatímco Degnan & Rosenberg (2006) ukázali, že v anomální zóně budou upřednostňovány symetrické genové stromy, i když je "pravý" (druhový) strom asymetrický, autoři zaznamenali jediný symetrický genový strom. Dokonce i po vyloučení asymetrických stromů s nepštrosími paleognáty se ve zbylém vzorku stále nejčastěji vyskytuje asymetrický strom, na němž jsou sesterskou skupinou všech ostatních paleognátů nanduové (což opět implikuje "běžčí" polyfylii). Zdá se tedy, že v rozporu se závěry Bakera a Haddratha (2010) nonmonofylie "běžců" není artefaktem nekompletního třídění linií a následných konfliktů mezi genovými stromy a druhovým stromem. Přídavné rozbory dále ukázaly, že monofyletičtí nepštrosí paleognáti jsou stejnou měrou podporováni anonymními markery a intronovými sekvencemi a že mezi lokusy podporujícími a odporujícími "běžčí" polyfylii nejsou žádné podstatné rozdíly v zastoupení jednotlivých bází. Klad nepštrosích paleognátů je tedy podpořen silným signálem rozloženým po celé šíři genomu.

3. Kam s tinamami
Mohlo by se zdát, že fylogenomika už všechny otázky paleognátí fylogeneze vyřešila: "běžce" musíme zavrhnout, nejbazálnějšími žijícími paleognáty jsou ve skutečnosti pštrosi a kiviové nestojí nějak zvlášť blízko báze paleognátů, ale hned vedle odvozených kasuárovitých (Hackett et al. 2008; Harshman et al. 2008; Smith et al. 2012). Víme dokonce víc, než bychom se odvažovali doufat: nejintenzivnější výzkum v oblasti studia pravěké DNA (tzv. "ancient DNA", aDNA) se nyní zaměřuje na ptáky moa (Allentoft & Rawlence 2011), kde se podařilo získat sekvence od stovek různých exemplářů a od kompletních mitochondriálních genomů se skrze mikrosatelity pomalu, ale jistě přesouváme k jadernému materiálu. MtDNA nejenže rozřešila vzájemné vztahy mezi jejich jednotlivými taxony (Bunce et al. 2009), ale pomohla tyto taxony i vymezit – v čemž selhala morfologie, neschopná odlišit geografickou variaci a pohlavní dimorfizmus od informativních rozdílů (viz revizi v Allentoft & Rawlence 2011) – a ukázala, že moa jsou sesterskou skupinou tinam (Phillips et al. 2010). Tento vztah zatím podpořila jediná studie, ale setkal se buď přímo s rychlým přijetím (Allentoft & Rawlence 2011), nebo neutrální reakcí ve stylu "bude potřeba víc dat" (Mayr 2011). Worthy & Scofield (2012) jej sice v dosud největší provedené morfologické analýze paleognátů neodkryli a vrátili se k hypotéze o monofyletických "běžcích", aspoň ale nepotvrdili ani nejzávažnější konkurenční hypotézu, totiž sesterský vztah moa a kiviů, a jejich pokus podat výsledky molekulárních studií jako nejednozančné vyzněl poněkud zoufale.
Technicky vzato tak není vůbec těžké otázku z nadpisu zodpovědět: tinamy pravděpodobně patří hned vedle ptáků moa. Ovšem na problému, kam s výsledným příbuzenským svazkem dál, si vylámaly zuby desítky jaderných lokusů i morfologických znaků a jde jednoznačně o nejzávažnější (a do jisté míry i poslední skutečný zbývající) problém ve fylogenetice žijících paleognátních ptáků. V rámci nepštrosích paleognátů, jejichž monofylii už můžeme bezpečně předpokládat (viz výše), rozeznáváme mimo tinam dvě další monofyletické skupiny. Jsou to nanduové (Rheidae nebo Rhea) a australasijští běžci, mezi jejichž žijící zástupce patří kiviové (Apteryx), emu (Dromaius) a kasuáři (Casuarius); pravděpodobně sem však patří i sloní ptáci (McDowell 1948*; Cooper et al. 2001; Phillips et al. 2010). Pro tři taxony lze sestrojit tři zakořeněné stromy, a právě tolika způsoby můžeme umístit tinamy na paleognátí strom – a pro každý z nich existuje nějaká podpora. Tinamy by mohly být sesterskou skupinou k australasijským běžcům a nanduové stát mimo (Chojnowski et al. 2008; Hackett et al. 2008; Harshman et al. 2008: věrohodnostní analýza s normálním kódováním, bayesovská analýza; Phillips et al. 2010), nebo by australasijští běžci mohli tvořit sesterský taxon ke kladu složenému z tinam a nanduů (Harshman et al. 2008: věrohodnost s RY kódováním, úspornostní analýza), a nebo by se tinamy mohly nacházet zcela mimo skupinu tvořenou nanduy a australasijskými běžci (Johnston 2011). Horší je, že 40 nových lokusů s celým problémem prakticky vůbec nepomáhá.
*Je to překvapivé, ale McDowellova studie, která si získala takovou oblibu mezi zastánci paleognátí polyfylie, předkládá silné důkazy pro sjednocení sloních ptáků s kivii, tedy přesně tentýž vztah, který vyhodilo i několik stovek bází mtDNA pracně vydolovaných ze subfosilního tibiotarzu o více než půl století později. Oba taxony sdílí tvar i pozici bazipterygoidních výběžků, kost patrovou ve tvaru silně zakřiveného tenoučkého plátu a rozdvojený přední konec křídlové kosti. McDowell si nebyl svazkem obou taxonů na základě svých pozorování příliš jist, ale jejich shoda s novou molekulární evidencí je pozoruhodná.
Nad rozšířeným taxon samplingem (tj. včetně kiviů, kasuára, ptáků moa atd.):
Naději, která zdánlivě svítá z odlišné citlivosti obou hypotéz na alignment, rychle zabíjí analýza v BUCKy. Ta sice opět vykrývá klad emu/tinamy vylučující nanduy, ale zároveň ukazuje, že ho podporuje jen 51% celého datasetu, přičemž 95% interval kredibility je mimořádně široký (32–71%) a podpora pro klad nanduové/tinamy vylučující emu není o moc nižší (44%, interval kredibility 25–64%). Co je nejhorší, oba intervaly kredibility zahrnují i hodnotu, kterou bychom očekávali od tvrdé polytomie (hard polytomy) – tento pojem popisuje situaci, kdy se z jednoho uzlu odštěpují více než dvě větve nikoli proto, že by analýza neměla dost dat na to, aby pořadí jejich odštěpování určila, ale protože se to tak prostě opravdu stalo: z mateřské populace se simultánně oddělily více než dvě dceřinné vývojové větve. S rozřešením tvrdé polytomie nepomůže sebevětší množství dat; pokud s přidáním dalších dat ze stromu nějaká polytomie zmizí, pak nebyla tvrdá. Prakticky každý fylogenetický problém, který systematiky už opravdu dlouho otravuje, byl jednou či vícekrát označen za tvrdou polytomii; v případě ptáků to udělali Poe & Chubb (2004), podle nichž takové charakteristiky vykazuje báze kladu Neoaves (ten zahrnuje všechny ptáky mimo paleognátů, hrabavých a vrubozobých). Chojnowski et al. (2008) tento návrh vyvrátili, když ukázali, že genové stromy několika různých lokusů se navzájem shodují více, než bychom pod hypotézou tvrdé polytomie očekávali – pak by totiž byla jediným zdrojem shody náhoda. Nyní se Smith et al. (2012) paradoxně obávají toho, že Chojnowski a spol. přehlédli úplně jinou tvrdou polytomii, totiž tu u báze nepštrosích paleognátů. Nebylo by to nemožné. Délky obou problematických větví (tj. větve sjednocující tinamy s nanduy a větve sjednocující tinamy s australasijskými běžci) jsou extrémně krátké, což by odpovídalo: tvrdá polytomie má příslušné délky větví nulové.

Smith a spol. zde na chvíli odbočují a pokoušejí se najít jiná kritéria, podle nichž by šlo mezi alternativními hypotézami rozhodnout. Maxwell & Larsson (2009) jsou citováni jako autoři pozorování, podle nějž tinamy a emuové sdílejí ztrátu dvou konkrétních prstních článků v křídle, kterými nanduové a pštrosi stále ještě disponují. To je zajímavé (a slyším o tom poprvé, takže i příjemné překvapení), ale nic moc to neznamená. Zaprvé: pokus rekonstruovat fylogenezi paleognátů ze znaků týkajících se redukovaných předních končetin tady už byl (Bledsoe 1988), a i když dopadl zajímavě – až do analýz Grellet-Tinnera a Johnstona šlo o jedinou morfologickou studii, která vykryla sesterský vztah kiviů a kasuárovitých –, Mayr (2011) uvádí, že byl právě z hlediska volby znaků kritizován. Zadruhé, morfologických důkazů pro klad složený z tinam a nanduů je ještě daleko více. Smith a spol. sice žádné z ních necitují, ale ve starší literatuře jich lze najít hodně, dokonce tak hodně, že opět vyvolávají dojem sekundárního signálu skrytého pod konvergencemi odrážejícími nelétavost a neotenii (viz níže). Autoři naopak zmiňují, že seskupení tinam a nanduů je daleko přijatelnější z hlediska biogeografie: obě skupiny se vyskytují pouze v Jižní Americe, odkud žádného jiného žijícího paleognáta neznáme. Zároveň ale ihned uvádějí problémy tohoto argumentu: i kdyby nanduové měli k tinamám blíže než australasijští běžci, pořád k nim mají ještě blíž rovněž australasijští ptáci moa. Dodal bych ještě, že další problém vyvstává u slůvka "žijícího". Má-li Alvarenga (2010) pravdu a Diogenornis ze středního paleocénu Brazílie skutečně patří do kmenové linie kasuárovitých spíše než nanduovitých (contra Mayr 2009), pak se biogeografický argument bortí ještě důkladněji a interpretovat tinamy a nanduy jako dvě nezávislé invaze do Jižní Ameriky z Australásie by bylo přinejmenším stejně úsporné jako vydávat ptáky moa za nezávislou invazi do Australásie z Jižní Ameriky.
Přesto je však závěr optimistický. Nepštrosí paleognáti nejspíše tvrdá polytomie nejsou, a to z jednoho prostého důvodu: ačkoli zatím nelze rozhodnout mezi sesterským vztahem tinam s australasijskými běžci a tinam s nanduy, vyloučit třetí hypotézu (tj. klad nanduů a australasijských běžců bez tinam) je až překvapivě snadné. Ve skutečnosti je podivné, jak tvrdošíjně ji data zamítají: pokud podrobíme každý z 40 lokusů samostatné bayesovské analýze a prozkoumáme obsah 95% kredibilní množiny stromů, ani v jediném případě v ní neobjevíme topologii (tinamy, (nanduové, emu)). Něco takového je s modelem tvrdé polytomie zcela neslučitelné. Špatnou zprávou je, že ačkoli je tato polytomie měkká, ani 61 lokusů s ní nepohne. Kolik s ní pohne? Těžko říct, ale autoři v této souvislosti citují nedávnou studii zaměřenou na ultrakonzervované elementy, která měla lokusů 854 (Faircloth et al. 2012).
4. Svět bez "běžců"
V diskuzi o biogeografických a evolučních implikacích své studie se Smith et al. (2012) nepřekvapivě obhajují vícenásobnou ztrátu letu u paleognátů. Alternativní možnost, tedy jedinou ztrátu letu u společného předka paleognátů, následovanou jeho znovuzískáním u tinam, považují za nepravděpodobnou. Zatímco ptáci ztrácí let velice snadno (stalo se to hned několika taxonům nejisté fylogenetické pozice – sylviornisovi, gastornitidům a dromornitidům; vymřelým kachnám moa-nalo, drontemu, tučňákům, vymřelému ibisovi Xenicibis, alce velké, forusracidům, kakapovi, vymřelé sově Ornimegalonyx, a počet "druhů" nelétavých chřástalů se blíží dvěma tisícům [Steadman 1995]), ztráta letu a jeho opětovné nabytí nebylo zaznamenáno nikdy. Dokonce i u hmyzu je jediný navržený případ poměrně kontroverzní (Whiting et al. 2003). A pokud různé skupiny "běžců" ztratily let nezávisle na sobě, znamená to, že nezávisle vznikly i některé jejich nejtypičtější znaky. Tato konvergence je daleko patrnější v postkraniu; proto se také jediné morfologické studie podporující "běžčí" polyfylii soustředily na lebku.
Polyfylie "běžců" také dává daleko větší volnost hypotézám vysvětlujícím jejich výskyt. Není už důvod spojovat fylogenetické divergence s rozpadem Gondwany – žádná z fylogenetických hypotéz vyjma té Johnstonovy (2011) mu stejně zcela neodpovídá, a ta Johnstonova úplné konzistence dosahuje jen za cenu rozporů s molekulárními studiemi v otázkách pozice tinam, moa a sloních ptáků. Molekulární data silně podporují hypotézu, že Nový Zéland byl paleognáty osídlen dvakrát nezávisle na sobě: jednak předky dnešních kiviů, jednak předky ptáků moa. Přestože fylogeneze sama neumožňuje vyloučit možnost, že "běžčí" evoluci tvaroval kontinentální drift, existují dva druhy důkazů svědčící o opaku.
Kdyby tomu tak bylo, musela by se velká část paleognátí evoluce odehrát v druhohorách, neboť na rozhraní křídy a paleogénu už byly kontinenty víceméně na svých místech. Pak je ale podezřelé, že téměř všechny paleognátí fosilie známe až z kenozoika. Hope (2002) ukázala, že většina mezozoických ptáků pokládaných aspoň čas od času za paleognáty buďto nejsou ptáci (Wyleyia), nejsou součástí ptačího korunního kladu (Ambiortus, Gansus), nebo jejich paleognátí status není možné přesvědčivě podpořit (Palaeocursornis). Nejstarší paleognátí fosilie, "litornitidi" ze souvrství Hornerstown (Parris & Hope 2002), pocházejí buď z nejpozdnější křídy, nebo nejstaršího paleogénu. Přítomnost odvozených vrubozových typu vegavise ve svrchnokřídových vrstvách a také všechny studie používající molekulární datování jednoznačně říkají, že nějací paleognáti v druhohorách být museli. Jak ale podotkli Houde & Haubold (1987), pokud by to skutečně měli být velcí "běžci" dnešního typu, pak bychom už nějaké měli znát, protože jejich velké, masivní, nepneumatizované kosti jsou při fosilizaci oproti pozůstatkům létajících ptáků ve výhodě. Krom toho známe z paleogénu Evropy a Severní Ameriky řadu paleognátů, nelétavých (Palaeotis) i létavých (Lithornithidae), kteří se sem museli nějak dostat. Smith et al. (2012) navrhují, že hypotéza o aktivním rozptylu přes oceán kombinovaném s několika oddělenými ztrátami letu dovede tento fakt vysvětlit lépe než jediný vznik nelétavosti s následnou vikariancí, který preferují zastánci "běžčí" monofylie. Odkazují na Houde'a (1988), podle nějž byli litornitidi zdatní letci schopní překonat oceán. Druhým důkazem pro to, že evoluce paleognátů s rozpadem Gondwany příliš nesouvisela, jsou podle Smithe a spol. molekulární hodiny, které ukazují, že data divergencí v rámci paleognátů jsou podstatně mladší než data separací jednotlivých pevnin z mateřského superkontinentu.
Zdá se tedy, že nám všem, kdo jsme si užívali "epyornisovu povídku" z Dawkinsova Příběhu předka (Dawkins 2004), zbydou jenom oči pro pláč. (Je to mimochodem už podruhé, co v článku do téhle knížky rýpu. Měl bych své oblíbené čtivo roztrhat a spálit?) Skvěle to pojmenovali Harshman s kolektivem: představa "běžců" postupně se vzdalujících svým příbuzným tak, jak je na obrovských "vorech" unáší nemilosrdný kontinentální drift,
- Proč neexistují "běžci" (Ratitae), díl 1. – co DNA skrývala a morfologové přehlíželi
- Proč neexistují "běžci" (Ratitae), díl 3. – protože to říkají retropozony
V předchozím díle minisérie jsem ukázal, jak se zrodila hypotéza o "běžcích" coby polyfyletickém seskupení ptáků v rámci monofyletické skupiny Palaeognathae, a vyjmenoval jsem nedlouhou řádku studií, které tento závěr zatím podpořily. Zatím jsem ale neukázal důvod, proč by tento scénář měl být důvěryhodnější než všechny alternativy, zvláště když morfologické i molekulární studie pro "běžčí" monofylii v předchozích dekádách snesly tolik důkazů. To by měl článek níže napravit.

Byl jsem v zoo! Vlevo nandu Darwinův (Rhea/Pterocnemia pennata), vpravo emu hnědý (Dromaius novaehollandiae). Souboj se bude vést o fylogenetickou blízkost k tinamám; jako zbraně si oba soupeři volí 40 jaderných genů, mtDNA, biogeografii a zapadlé morfologické studie (Zdroj: autorova fotografie)
1. Smith et al. 2012: fylogenetika, metody a problémy
Skupina autorů z týmu Early Bird se rozhodla ověřit otázku asi ze všech nejdůležitějších, totiž jak dobře si vlastně hypotéza "běžčí" polyfylie stojí. Důvodů k pochybám je několik. Zaprvé existuje několik studií jaderných genů, které monofylii skupiny "Ratitae" podpořily (např. již citovaní García-Moreno & Mindell 2000). Zadruhé jednotlivé "běžce" stále spojuje velké množství odvozených morfologických znaků, především pak z postkrania, a ačkoli je lákavé odvysvětlit je neotenií, nikdy nebylo prokázáno, že neotenie dokáže v případě "běžců" vyprodukovat dostatečně mnoho konvergencí k tomu, aby zkreslovala výsledky fylogenetických analýz. Zatřetí ani datasety tak velké, jako jsou ty sestavené týmem Early Bird (Hackett et al. 2008; Harshman et al. 2008), nejsou zárukou dosažení správných výsledků. Např. postavení žebernatek v rámci metazoí je případem, kdy i velmi velké datasety podávají rozporuplné výsledky. Zvětšování datasetu je dobrou strategií pro eliminaci náhodného šumu, ale v molekulární evoluci je zdokumentováno mnoho jevů, které mohou výsledky analýzy zkreslit nenáhodně, tj. určitým směrem. Dvěma takovými biasy jsou přitahování dlouhých větví (long branch attraction) a konvergence na podobném zastoupení bází v sekvenci (base composition bias) a právě pomocí nich zastánci "běžčí" polyfylie vysvětlují, proč monofyletičtí "běžci" vycházeli i z tolika molekulárních studií: tinamy jsou dlouhá větev a přitahují se k outgroup, pštrosi zase sdílejí nozký obsah tyminu s cytozinem v mitochondriálním genomu s kivii a kasuárovitými.
Krom těchto jevů ale může fylogenezi zkreslit i nesoulad mezi genovými stromy (gene trees) a druhovými stromy (species trees) v důsledku nekompletního třídění linií. Smith et al. (2012) je sice necitují, ale Baker & Haddrath (2010) v abstraktu z předloňského ornitologického kongresu navrhli, že by "běžčí" polyfylie mohla být artefaktem právě tohoto jevu. Na tomtéž místě zmiňují, že když na rozsáhlý, dosud nepublikovaný dataset aplikovali pokročilé bayesovské metody pro rekonstrukci druhového stromu místo prostého zřetězení (concatenation) sekvencí jednotlivých genů, vyšli jim "běžci" zase monofyletičtí. A s tím, jak poznáváme lépe a lépe zákonitosti genomové evoluce, nacházíme nové a nové problémy. Co třeba heterotachie, tedy jev, při němž se rychlost substituce na jediné nukleotidové pozici mění v čase (a tedy napříč fylogenetickým stromem)? Nechceme-li se vzdát výhod parametrických metod, musíme využít modelů, které věrohodnost pro každý znak počítají jako vážený součet přes několik množin délek větví (mixture models). Jenže Matsen & Steel (2007) ukázali, že analýza dvou genů, jejichž fylogenetické stromy se liší pouze v délkách větví (tj. topologie genových stromů je stejná), nám pod "mixture model" může vyhodit strom s topologií úplně odlišnou. Problémy jsou i s alignmentem: fylogenetická analýza špatně zarovnaných sekvencí vyhodí špatný strom. A nemělo by se zapomínat ani na to, že podpora jednotlivých uzlů je skoro důležitější než samotný bodový odhad stromu, takže teoretičtí fylogenetici se vyžívají v hledání situací, kdy naprosto chybné klady obdrží vysoké hodnoty bootstrapu a posteriorních pravděpodobností. Co když je drasticky nadhodnocena např. vysoká podpora pro nepštrosí paleognáty v analýzách Harshmana a spol.?
Smith a spol. proto otestovali hypotézu "běžčí" polyfylie jednou z nejjednodušších metod vůbec: vzali nezávislá data (tj. ta, která nebyla zahrnuta v žádné z analýz, která původní hypotézu podpořila) a podívali se, co z nich vyleze. Podobné testování kontroverzních hypotéz má v ptačí systematice tradici: Morgan-Richards et al. (2008) se takto podívali na zoubek kladu Metaves skrze mitochondriální data (a hypotézu zamítli), Wang et al. (2011) loni zase s 30 novými jadernými geny ověřovali existenci skupiny Eufalconimorphae, složené z pěvců, papoušků a sokolů (hypotézu v plném rozsahu potvrdili, ale ukázali, jak zoufalá je situace v případě myšáků). Smith a spol. jsou ještě o něco ambicióznější: předkládají hned 40 zbrusu nových lokusů roztroušených po celém jaderném genomu. Dohromady dávají přes 22 000 bází, což konkuruje původnímu datasetu Harshmana a spol., a lze mezi nimi najít i "anonymní lokusy", tzn. náhodné fragmenty genomu, které mohou zahrnovat i dlouhé introny a mezigenové regiony. Pokud by tyto oblasti genomu nesly jiný signál než exony, krátké introny a UTRs použité Harshmanem a spol., Smith et al. (2012) by to dokázali zjistit.
Zarovnávání sekvencí Smith a spol. provedli manuálně ("eyeball alignment"), přičemž problematické sekvence bez milosti vyřazovali. Zároveň ale vyzkoušeli i trochu jiný postup: aby prověřili, jak moc je analýza na zarovnání citlivá, nakrmili program PRANK (provádějící alignment automaticky na základě maximální věrohodnosti) 15 tzv. "guide trees". Jelikož se PRANK snaží určit, za kterou mezeru (gap) v zarovnané sekvenci je zodpovědná delece a za jakou inzerce, potřebuje strom – "guide tree" –, na němž by mohl zrekonstruovat ancestrální sekvence. Když se mu žádný nezadá, PRANK si ho sestaví sám skrze neighbor-joining, ale Smith et al. (2012) této možnosti nevyužili. Místo toho mu postupně zadali všechny stromy, které uznali za realistické: "realističnost" přitom předpokládá monofylii nanduů (Rhea + Pterocnemia), monofylii tinam (Tinamus + Crypturellus) a pevně fixovanou topologii outgroup (která předpokládá monofylii hrabavých a drůbeže, čili opět nic kontroverzního). Proměnlivé byly jen vztahy pštrosů, nanduů, tinam a emu, což dá výše zmíněných 15 stromů. Délky jejich větví autoři spočítali pomocí věrohodnostní analýzy nad manuálním alignmentem.
Pokud jde o hlavní analýzu, byli Smith a spol. velice opatrní. Konstatují, že obrovské mnohalokusové datasety sice slibují mnohé, ale jen tehdy, pokud nespadají do problematické oblasti parametrového prostoru: narážejí na "anomální zónu", kterou popsali Degnan & Rosenberg (2006) a která má potenciál být fylogenetiky vyslovována se stejným nádechem zlověstnosti, jenž už léta doprovází zónu Felsensteinovu. V této zóně mají druhové stromy takovou kombinaci krátkých vnitřních větví, která vygeneruje genové stromy odporující stromu druhovému častěji než ty, které se s druhovým stromem shodují. Kubatko & Degnan (2007) pak ukázali, že k tomu, aby se maximální věrohodnost stala při rekonstrukci druhového stromu ze zřetězených sekvencí statisticky nekonzistentní, se stačí k obávané zóně jen přiblížit. Smith s kolektivem tedy proto kromě zřetězeného datasetu analyzovali metodami nejvyšší parsimonie a věrohodnosti i každý lokus zvlášť, přičemž věrohodnostní analýzu ještě provedli jak s jedním modelem naraženým na celý dataset, tak i se samostatným modelem pro každý lokus. Trojku hlavních fylogenetických metod doplnila bayesovská inference, pod níž lze navíc nasadit modely adresující výše zmíněný problém heterotachie. Smith et al. (2012) vyzkoušeli hned dva: jednak "mixture model" popsaný výše, jednak "covarion model", který se heterotachii pokouší obelstít trochu jinak: používá skrytý Markovův proces*, který dovoluje, aby se nukleotidové pozice během své evoluce přepínaly ze stavu "off" (kdy na nich žádné substituce neprobíhají) do stavu "on", kdy k mutacím dochází normálně (Kolaczkowski & Thornton 2008). Tato celkem jednoduchá původní myšlenka Tuffleyho a Steela (1998) byla časem doplněna řadou rafinovaných vymyšleností, takže původní binární volba "on/off" byla rozšířena na výběr z vícero rychlosntních kategorií (takový model vlastně dělá s variací v tempu substituce napříč časem totéž, co klasický gama parametr s variací v tempu substituce napříč nukleotidovými pozicemi) a posléze i kombinaci obou předchozích možností.
*To, že je Markovův, znamená, že pravděpodobnost další události nezávisí na událostech předchozích. V jazyce molekulární evoluce to znamená, že důležité je jen to, jaký je stav nukleotidové pozice teď, ne to, jaký byl předtím. To, že je skrytý, znamená, že jeho jednotlivé stavy nemůžeme přímo pozorovat – tyto "skryté" stavy jsou ale nějakým mechanizmem propojeny s pozorovatelnými výstupy a lze je z nich odhadnout.
Rozpory mezi genovými a druhovými stromy Smith a spol. zjišťovali pomocí programu BUCKy, který bayesovsky (dvojnásobnou aplikací Markovových řetězců Monte Carlo) odhaduje tzv. faktory konkordance (shody), tedy procento genomu, které podporuje zadaný klad. Jako každá bayesovská metoda, i BUCKy vyžaduje specifikování priorní pravděpodobnosti: zde jde o parametr α, který vyjadřuje apriorní přesvědčení systematika o tom, jak moc si budou jednotlivé genové stromy z jeho datasetu navzájem odporovat (α = 0: vůbec, α = ∞: historie jednotlivých genů budou absolutně nezávislé). S priorními pravděpodobnostmi je vždycky trochu potíž: ve fylogenetice třeba panuje shoda na tom, že všechny topologie by měly mít stejný prior – žádný strom bychom neměli upřednostňovat dřív, než analyzujeme data. Podstatně kontroverznější je volba priorů pro délky větví, a bayesovsky založení biologové si stěžují, že největší výzkum v oblasti priorních pravděpodobností se soustředí na to, jak je učinit neinformativními (Ronquist & Deans 2010). Ze Smithe a jeho spoluautorů by ale Ronquist s Deansem měli radost: jejich alfa je informativní, odvozená z předchozích studií paleognátí fylogeneze (Harshman et al. 2008; Phillips et al. 2010) a poskytující následující roztomilé priorní rozdělení pro počet odlišných genových stromů:

Program BUCKy má skvělý web, který z počtu genů, počtu taxonů a hodnoty parametru α spočítá priorní rozdělení pro počet genových stromů s odlišnými topologiemi. Smith et al. (2012) na základě předchozích studií nastavili alfu na 3, což spolu s použitými 40 lokusy a 6 ingroup taxony (emu, pštros, 2 tinamy, 2 nanduové) vyhodí toto rozdělení, podle nějž máme očekávat něco čtyřmi a dvanácti rozpornými stromy. Vytvořeno pomocí webové podpory pro BUCKy.

Totéž, tentokrát pro 4 taxony (vnitřní diverzita nanduů a tinam zanedbána). Počet očekávaných genových stromů najednou klesá na 3 – práve tolik topologií nakonec skutečně mělo nenulovou posteriorní pravděpodobnost.
Autoři použili BUCKy ke dvěma účelům: jednak aby určili, kolik lokusů podporuje ten který klad, jednak k rekonstrukci druhového stromu pomocí "kvartetové metody", která má tu příjemnou vlastnost, že je statisticky konzistentní a lze garantovat, že správnou topologii druhového stromu najde i tehdy, když neodpovídá nejčastěji pozorovanému genovému stromu. Vyhýbá se tak výše zmíněné anomální zóně a bere v úvahu fakt, že postup, který Dawkins (2004) popsal jako "demokratické hlasování" jednotlivých genů, může být při rekonstrukci evoluční historie organizmů a populací zavádějící. Nevýhodou je pak skutečnost, že statistické konzistence může dosáhnout jen tehdy, když jsou faktory konkordance odhadnuty správně. Autoři ale dokázali obejít i tento požadavek. Rozdělili genové stromy do dvou kategorií – na ty, na nichž klad všech nepštrosích paleognátů je, a na ty, na kterých není – a pak spočítali, kolik stromů by v obou kategorií mělo být, pokud by klad nepštrosích paleognátů neexistoval (na druhovém stromě). Následně použili statistický test, aby zjistili, zda lze tuto nulovou hypotézu zamítnout.
Jako tečku za záplavou dílčích analýz Smith et al. (2012) zkombinovali svá data se sekvencemi Harshmana a spol. (2008) a Phillipse a spol. (2010), čímž složili supermatici atakující 60 kilobází. Podobný postup zvolili Wang et al. (2011) při loňském přetestování sesterského vztahu mezi pěvci a papoušky: ať už spolu jednotlivé datasety budou souhlasit nebo ne, nejrobustnějších výsledků dosáhneme, když je spojíme dohromady. Analýzy provedené nad tímto datasetem byly celkem dvě: jedna s minimálním taxon samplingem, tj. pouze s taxony společnými pro všechny dílčí datasety (pštrosa, dvě tinamy, oba nanduy a emu), druhá měla naopak taxon sampling maximální, zahrnující všechny taxony přítomné aspoň v jednom datasetu – celkem tedy pštrosa, oba nanduy, čtyři tinamy, tři ptáky moa, tři kivie, emu a kasuára. Dva kiviové a všichni moa byli v analýze reprezentováni výhradně mitochondriálními sekvencemi. Na oba tyto datasety byla nasazena parsimonie, věrohodnost i bayesovská inference; data byla opět roztříděna do několika oddílů se zvláštním modelem pro každý jeden jaderný gen. V detailech analýzy mitochondriální části datasetu Smith s kolektivem následovali to, co zvolili Phillips et al. (2010).
2. "Běžci" verzus nepštrosí paleognáti: souboj hypotéz na život a na smrt
Ve stručnosti: klad nepštrosích paleognátů (a tím pádem polyfylie "běžců") byl jednoznačně potvrzen. Nejprve autoři prozkoumali obsah 95% kredibilní množiny stromů (95% credible set of trees). Tuto množinu dostaneme tak, že ve vzorku, který sestavily Markovovy řetězce Monte Carlo (MCMC) v průběhu bayesovské analýzy, seřadíme jednotlivé stromy podle jejich posteriorní pravděpodobnosti (PP). Postupujeme od stromu s nejvyšší posteriorní pravděpodobností k méně a méně pravděpodobným stromům, dokud celková posteriorní pravděpodobnost nedosáhne hodnoty 0,95. Asi se do ní netrefíme úplně přesně (když má 1. strom PP 0,45, druhý 0,35 a třetí 0,18, celkový součet je už 0,98), ale jak ve své výborné studii o interpreaci bayesovských pojmů a měřítek ukázali Huelsenbeck & Rannala (2004), to lze napravit tím, když strom překračující 95% hranici zahrneme jen s pravděpodobností odpovídající tomu, o kolik ji překračuje. Ukázalo se, že v 95% kredibilní množině stromů se nachází jen jediná topologie, a ta obsahuje nepštrosí paleognáty. Topologie, na nichž se vyskytují monofyletičtí "běžci", byly vyloučeny dokonce z 99% kredibilní množiny stromů: ač tvoří 3 z 15 (tj. 20%) možných zakořeněných stromů popsaných výše, na posteriorní pravděpodobnosti se podílejí méně než 1%.
"Běžčí" polyfylii ve výsledcích určitě nelze svést na heterotachii, tedy změny v rychlosti evoluce za čas. Bayesovská analýza s využitím "covarion" a "mixture" modelů evoluce odhalila monofyletické nepštrosí paleognáty zase, a to s posteriorní pravděpodobností 1. Ukázalo se navíc, že MCMC pro většinu větví nezvolily víc než jednu délku (ačkoli klidně mohly). Rozbor jednotlivých genových stromů ukázal, že 24 z 38 lokusů podporuje monofylii nepštrosích paleognátů ve věrohodnostní i bayesovské analýze, zatímco pod úspornostní analýzou je to 15 z 38 (chybějící 2 lokusy nemohly být osekvenovány od jednoho z paleognátích taxonů, a tak je autoři pro teď ignorovali) – v obou případech je to víc než 16% a méně procent, které bychom očekávali, kdyby takový klad nebyl přítomen na druhovém stromě. Podle autorů může menší podpora ze strany parsimonie odrážet náchylnost metody k přitahování dlouhých větví, zde tedy konkrétně dlouhé větve tinam k dlouhé větvi outgroup. Faktor konkordance ukazuje, že monofylii nepštrosích paleognátů podporuje 93% genomu – 95% interval kredibility (tj. interval, který s pravděpodobností 95% obsahuje správnou hodnotu) je 84% až 99%. A zatímco Degnan & Rosenberg (2006) ukázali, že v anomální zóně budou upřednostňovány symetrické genové stromy, i když je "pravý" (druhový) strom asymetrický, autoři zaznamenali jediný symetrický genový strom. Dokonce i po vyloučení asymetrických stromů s nepštrosími paleognáty se ve zbylém vzorku stále nejčastěji vyskytuje asymetrický strom, na němž jsou sesterskou skupinou všech ostatních paleognátů nanduové (což opět implikuje "běžčí" polyfylii). Zdá se tedy, že v rozporu se závěry Bakera a Haddratha (2010) nonmonofylie "běžců" není artefaktem nekompletního třídění linií a následných konfliktů mezi genovými stromy a druhovým stromem. Přídavné rozbory dále ukázaly, že monofyletičtí nepštrosí paleognáti jsou stejnou měrou podporováni anonymními markery a intronovými sekvencemi a že mezi lokusy podporujícími a odporujícími "běžčí" polyfylii nejsou žádné podstatné rozdíly v zastoupení jednotlivých bází. Klad nepštrosích paleognátů je tedy podpořen silným signálem rozloženým po celé šíři genomu.

Fylogenetický strom vzešlý z analýzy 40 lokusů v programu BUCKy, navrženém ke statisticky konzistentnímu odhadování druhových stromů ze vzájemně rozporných stromů genových. Čísla nad větvemi udávají, jaká část genomu podporuje existenci dané větve, čísla dole pak pro tento údaj poskytují 95% interval kredibility. Je vidět, že zatímco monofylii nepštrosích paleognátů podporuje stejné procento genomu jako monofylii hrabavých (větev Gallus + Crax) nebo kladu Galloanserae, tedy skupin zcela nekontroverzních, podpora pro větev sjednocující emu a tinamy je velmi slabá. Šedý obdélník vyznačuje taxony historicky řazené mezi "běžce" (Zdroj: Smith et al. 2012: Figure 3)
3. Kam s tinamami
Mohlo by se zdát, že fylogenomika už všechny otázky paleognátí fylogeneze vyřešila: "běžce" musíme zavrhnout, nejbazálnějšími žijícími paleognáty jsou ve skutečnosti pštrosi a kiviové nestojí nějak zvlášť blízko báze paleognátů, ale hned vedle odvozených kasuárovitých (Hackett et al. 2008; Harshman et al. 2008; Smith et al. 2012). Víme dokonce víc, než bychom se odvažovali doufat: nejintenzivnější výzkum v oblasti studia pravěké DNA (tzv. "ancient DNA", aDNA) se nyní zaměřuje na ptáky moa (Allentoft & Rawlence 2011), kde se podařilo získat sekvence od stovek různých exemplářů a od kompletních mitochondriálních genomů se skrze mikrosatelity pomalu, ale jistě přesouváme k jadernému materiálu. MtDNA nejenže rozřešila vzájemné vztahy mezi jejich jednotlivými taxony (Bunce et al. 2009), ale pomohla tyto taxony i vymezit – v čemž selhala morfologie, neschopná odlišit geografickou variaci a pohlavní dimorfizmus od informativních rozdílů (viz revizi v Allentoft & Rawlence 2011) – a ukázala, že moa jsou sesterskou skupinou tinam (Phillips et al. 2010). Tento vztah zatím podpořila jediná studie, ale setkal se buď přímo s rychlým přijetím (Allentoft & Rawlence 2011), nebo neutrální reakcí ve stylu "bude potřeba víc dat" (Mayr 2011). Worthy & Scofield (2012) jej sice v dosud největší provedené morfologické analýze paleognátů neodkryli a vrátili se k hypotéze o monofyletických "běžcích", aspoň ale nepotvrdili ani nejzávažnější konkurenční hypotézu, totiž sesterský vztah moa a kiviů, a jejich pokus podat výsledky molekulárních studií jako nejednozančné vyzněl poněkud zoufale.
Technicky vzato tak není vůbec těžké otázku z nadpisu zodpovědět: tinamy pravděpodobně patří hned vedle ptáků moa. Ovšem na problému, kam s výsledným příbuzenským svazkem dál, si vylámaly zuby desítky jaderných lokusů i morfologických znaků a jde jednoznačně o nejzávažnější (a do jisté míry i poslední skutečný zbývající) problém ve fylogenetice žijících paleognátních ptáků. V rámci nepštrosích paleognátů, jejichž monofylii už můžeme bezpečně předpokládat (viz výše), rozeznáváme mimo tinam dvě další monofyletické skupiny. Jsou to nanduové (Rheidae nebo Rhea) a australasijští běžci, mezi jejichž žijící zástupce patří kiviové (Apteryx), emu (Dromaius) a kasuáři (Casuarius); pravděpodobně sem však patří i sloní ptáci (McDowell 1948*; Cooper et al. 2001; Phillips et al. 2010). Pro tři taxony lze sestrojit tři zakořeněné stromy, a právě tolika způsoby můžeme umístit tinamy na paleognátí strom – a pro každý z nich existuje nějaká podpora. Tinamy by mohly být sesterskou skupinou k australasijským běžcům a nanduové stát mimo (Chojnowski et al. 2008; Hackett et al. 2008; Harshman et al. 2008: věrohodnostní analýza s normálním kódováním, bayesovská analýza; Phillips et al. 2010), nebo by australasijští běžci mohli tvořit sesterský taxon ke kladu složenému z tinam a nanduů (Harshman et al. 2008: věrohodnost s RY kódováním, úspornostní analýza), a nebo by se tinamy mohly nacházet zcela mimo skupinu tvořenou nanduy a australasijskými běžci (Johnston 2011). Horší je, že 40 nových lokusů s celým problémem prakticky vůbec nepomáhá.
*Je to překvapivé, ale McDowellova studie, která si získala takovou oblibu mezi zastánci paleognátí polyfylie, předkládá silné důkazy pro sjednocení sloních ptáků s kivii, tedy přesně tentýž vztah, který vyhodilo i několik stovek bází mtDNA pracně vydolovaných ze subfosilního tibiotarzu o více než půl století později. Oba taxony sdílí tvar i pozici bazipterygoidních výběžků, kost patrovou ve tvaru silně zakřiveného tenoučkého plátu a rozdvojený přední konec křídlové kosti. McDowell si nebyl svazkem obou taxonů na základě svých pozorování příliš jist, ale jejich shoda s novou molekulární evidencí je pozoruhodná.
Nad rozšířeným taxon samplingem (tj. včetně kiviů, kasuára, ptáků moa atd.):
- věrohodnostní (ML) analýza s jedním modelem evoluce pro celý zřetězený dataset spojuje klad moa/tinamy s nanduy;
- věrohodnostní (ML) analýza se samostatným modelem evoluce pro každý jeden lokus spojuje klad moa/tinamy s nanduy;
- bayesovská analýza spojuje klad moa/tinamy s australasijskými běžci;
- úspornostní (MP) analýza spojuje tinamy s nanduy;
- věrohodnostní (ML) analýza s jedním modelem evoluce pro celý zřetězený dataset spojuje tinamy s nanduy;
- věrohodnostní (ML) analýza se samostatným modelem evoluce pro každý jeden lokus spojuje tinamy s australasijskými běžci;
- bayesovská analýza se samostatným modelem evoluce pro každý jeden lokus spojuje tinamy s australasijskými běžci;
- druhový strom rekonstruovaný z genových stromů pomocí BUCKy spojuje tinamy s australasijskými běžci
Naději, která zdánlivě svítá z odlišné citlivosti obou hypotéz na alignment, rychle zabíjí analýza v BUCKy. Ta sice opět vykrývá klad emu/tinamy vylučující nanduy, ale zároveň ukazuje, že ho podporuje jen 51% celého datasetu, přičemž 95% interval kredibility je mimořádně široký (32–71%) a podpora pro klad nanduové/tinamy vylučující emu není o moc nižší (44%, interval kredibility 25–64%). Co je nejhorší, oba intervaly kredibility zahrnují i hodnotu, kterou bychom očekávali od tvrdé polytomie (hard polytomy) – tento pojem popisuje situaci, kdy se z jednoho uzlu odštěpují více než dvě větve nikoli proto, že by analýza neměla dost dat na to, aby pořadí jejich odštěpování určila, ale protože se to tak prostě opravdu stalo: z mateřské populace se simultánně oddělily více než dvě dceřinné vývojové větve. S rozřešením tvrdé polytomie nepomůže sebevětší množství dat; pokud s přidáním dalších dat ze stromu nějaká polytomie zmizí, pak nebyla tvrdá. Prakticky každý fylogenetický problém, který systematiky už opravdu dlouho otravuje, byl jednou či vícekrát označen za tvrdou polytomii; v případě ptáků to udělali Poe & Chubb (2004), podle nichž takové charakteristiky vykazuje báze kladu Neoaves (ten zahrnuje všechny ptáky mimo paleognátů, hrabavých a vrubozobých). Chojnowski et al. (2008) tento návrh vyvrátili, když ukázali, že genové stromy několika různých lokusů se navzájem shodují více, než bychom pod hypotézou tvrdé polytomie očekávali – pak by totiž byla jediným zdrojem shody náhoda. Nyní se Smith et al. (2012) paradoxně obávají toho, že Chojnowski a spol. přehlédli úplně jinou tvrdou polytomii, totiž tu u báze nepštrosích paleognátů. Nebylo by to nemožné. Délky obou problematických větví (tj. větve sjednocující tinamy s nanduy a větve sjednocující tinamy s australasijskými běžci) jsou extrémně krátké, což by odpovídalo: tvrdá polytomie má příslušné délky větví nulové.

Nejobsáhlejší z analýz provedených Smithem a spol. Použito bylo celkem 61 lokusů – 20 jaderných lokusů z analýzy Harshmana a spol. (2008), kompletní mitochondriální genomy ze studie Phillipse a spol. (2010) a 40 nových jaderných lokusů od Smithova týmu samotného. Zahrnuty rovněž byly všechny taxony použité alespoň v jedné z těchto tří analýz. Je patrné, že na výsledném stromě se opět objevil klad (tinamy + nanduové), i ten je však podložen velice slabě. Čísla kolem větví udávají v pořadí odshora dolů bootstrapové hodnoty pro: věrohodnostní analýzu dělenou do oddílů podle lokusů, věrohodnostní analýzu s jediným modelem evoluce pro celý zřetězený dataset, úspornostní analýzu; a hodnotu posteriorní pravděpodobnosti z bayesovské analýzy. "NP" znamená, že daný klad v příslušné analýze žádnou podporu nezískal. (Modifikováno ze Smith et al. 2012: Figure 4)
Smith a spol. zde na chvíli odbočují a pokoušejí se najít jiná kritéria, podle nichž by šlo mezi alternativními hypotézami rozhodnout. Maxwell & Larsson (2009) jsou citováni jako autoři pozorování, podle nějž tinamy a emuové sdílejí ztrátu dvou konkrétních prstních článků v křídle, kterými nanduové a pštrosi stále ještě disponují. To je zajímavé (a slyším o tom poprvé, takže i příjemné překvapení), ale nic moc to neznamená. Zaprvé: pokus rekonstruovat fylogenezi paleognátů ze znaků týkajících se redukovaných předních končetin tady už byl (Bledsoe 1988), a i když dopadl zajímavě – až do analýz Grellet-Tinnera a Johnstona šlo o jedinou morfologickou studii, která vykryla sesterský vztah kiviů a kasuárovitých –, Mayr (2011) uvádí, že byl právě z hlediska volby znaků kritizován. Zadruhé, morfologických důkazů pro klad složený z tinam a nanduů je ještě daleko více. Smith a spol. sice žádné z ních necitují, ale ve starší literatuře jich lze najít hodně, dokonce tak hodně, že opět vyvolávají dojem sekundárního signálu skrytého pod konvergencemi odrážejícími nelétavost a neotenii (viz níže). Autoři naopak zmiňují, že seskupení tinam a nanduů je daleko přijatelnější z hlediska biogeografie: obě skupiny se vyskytují pouze v Jižní Americe, odkud žádného jiného žijícího paleognáta neznáme. Zároveň ale ihned uvádějí problémy tohoto argumentu: i kdyby nanduové měli k tinamám blíže než australasijští běžci, pořád k nim mají ještě blíž rovněž australasijští ptáci moa. Dodal bych ještě, že další problém vyvstává u slůvka "žijícího". Má-li Alvarenga (2010) pravdu a Diogenornis ze středního paleocénu Brazílie skutečně patří do kmenové linie kasuárovitých spíše než nanduovitých (contra Mayr 2009), pak se biogeografický argument bortí ještě důkladněji a interpretovat tinamy a nanduy jako dvě nezávislé invaze do Jižní Ameriky z Australásie by bylo přinejmenším stejně úsporné jako vydávat ptáky moa za nezávislou invazi do Australásie z Jižní Ameriky.
Přesto je však závěr optimistický. Nepštrosí paleognáti nejspíše tvrdá polytomie nejsou, a to z jednoho prostého důvodu: ačkoli zatím nelze rozhodnout mezi sesterským vztahem tinam s australasijskými běžci a tinam s nanduy, vyloučit třetí hypotézu (tj. klad nanduů a australasijských běžců bez tinam) je až překvapivě snadné. Ve skutečnosti je podivné, jak tvrdošíjně ji data zamítají: pokud podrobíme každý z 40 lokusů samostatné bayesovské analýze a prozkoumáme obsah 95% kredibilní množiny stromů, ani v jediném případě v ní neobjevíme topologii (tinamy, (nanduové, emu)). Něco takového je s modelem tvrdé polytomie zcela neslučitelné. Špatnou zprávou je, že ačkoli je tato polytomie měkká, ani 61 lokusů s ní nepohne. Kolik s ní pohne? Těžko říct, ale autoři v této souvislosti citují nedávnou studii zaměřenou na ultrakonzervované elementy, která měla lokusů 854 (Faircloth et al. 2012).
4. Svět bez "běžců"
V diskuzi o biogeografických a evolučních implikacích své studie se Smith et al. (2012) nepřekvapivě obhajují vícenásobnou ztrátu letu u paleognátů. Alternativní možnost, tedy jedinou ztrátu letu u společného předka paleognátů, následovanou jeho znovuzískáním u tinam, považují za nepravděpodobnou. Zatímco ptáci ztrácí let velice snadno (stalo se to hned několika taxonům nejisté fylogenetické pozice – sylviornisovi, gastornitidům a dromornitidům; vymřelým kachnám moa-nalo, drontemu, tučňákům, vymřelému ibisovi Xenicibis, alce velké, forusracidům, kakapovi, vymřelé sově Ornimegalonyx, a počet "druhů" nelétavých chřástalů se blíží dvěma tisícům [Steadman 1995]), ztráta letu a jeho opětovné nabytí nebylo zaznamenáno nikdy. Dokonce i u hmyzu je jediný navržený případ poměrně kontroverzní (Whiting et al. 2003). A pokud různé skupiny "běžců" ztratily let nezávisle na sobě, znamená to, že nezávisle vznikly i některé jejich nejtypičtější znaky. Tato konvergence je daleko patrnější v postkraniu; proto se také jediné morfologické studie podporující "běžčí" polyfylii soustředily na lebku.
Polyfylie "běžců" také dává daleko větší volnost hypotézám vysvětlujícím jejich výskyt. Není už důvod spojovat fylogenetické divergence s rozpadem Gondwany – žádná z fylogenetických hypotéz vyjma té Johnstonovy (2011) mu stejně zcela neodpovídá, a ta Johnstonova úplné konzistence dosahuje jen za cenu rozporů s molekulárními studiemi v otázkách pozice tinam, moa a sloních ptáků. Molekulární data silně podporují hypotézu, že Nový Zéland byl paleognáty osídlen dvakrát nezávisle na sobě: jednak předky dnešních kiviů, jednak předky ptáků moa. Přestože fylogeneze sama neumožňuje vyloučit možnost, že "běžčí" evoluci tvaroval kontinentální drift, existují dva druhy důkazů svědčící o opaku.
Kdyby tomu tak bylo, musela by se velká část paleognátí evoluce odehrát v druhohorách, neboť na rozhraní křídy a paleogénu už byly kontinenty víceméně na svých místech. Pak je ale podezřelé, že téměř všechny paleognátí fosilie známe až z kenozoika. Hope (2002) ukázala, že většina mezozoických ptáků pokládaných aspoň čas od času za paleognáty buďto nejsou ptáci (Wyleyia), nejsou součástí ptačího korunního kladu (Ambiortus, Gansus), nebo jejich paleognátí status není možné přesvědčivě podpořit (Palaeocursornis). Nejstarší paleognátí fosilie, "litornitidi" ze souvrství Hornerstown (Parris & Hope 2002), pocházejí buď z nejpozdnější křídy, nebo nejstaršího paleogénu. Přítomnost odvozených vrubozových typu vegavise ve svrchnokřídových vrstvách a také všechny studie používající molekulární datování jednoznačně říkají, že nějací paleognáti v druhohorách být museli. Jak ale podotkli Houde & Haubold (1987), pokud by to skutečně měli být velcí "běžci" dnešního typu, pak bychom už nějaké měli znát, protože jejich velké, masivní, nepneumatizované kosti jsou při fosilizaci oproti pozůstatkům létajících ptáků ve výhodě. Krom toho známe z paleogénu Evropy a Severní Ameriky řadu paleognátů, nelétavých (Palaeotis) i létavých (Lithornithidae), kteří se sem museli nějak dostat. Smith et al. (2012) navrhují, že hypotéza o aktivním rozptylu přes oceán kombinovaném s několika oddělenými ztrátami letu dovede tento fakt vysvětlit lépe než jediný vznik nelétavosti s následnou vikariancí, který preferují zastánci "běžčí" monofylie. Odkazují na Houde'a (1988), podle nějž byli litornitidi zdatní letci schopní překonat oceán. Druhým důkazem pro to, že evoluce paleognátů s rozpadem Gondwany příliš nesouvisela, jsou podle Smithe a spol. molekulární hodiny, které ukazují, že data divergencí v rámci paleognátů jsou podstatně mladší než data separací jednotlivých pevnin z mateřského superkontinentu.
Zdá se tedy, že nám všem, kdo jsme si užívali "epyornisovu povídku" z Dawkinsova Příběhu předka (Dawkins 2004), zbydou jenom oči pro pláč. (Je to mimochodem už podruhé, co v článku do téhle knížky rýpu. Měl bych své oblíbené čtivo roztrhat a spálit?) Skvěle to pojmenovali Harshman s kolektivem: představa "běžců" postupně se vzdalujících svým příbuzným tak, jak je na obrovských "vorech" unáší nemilosrdný kontinentální drift,
should be viewed as yet another example of the phenomenon that Huxley called ‘‘the great tragedy of science—the slaying of a beautiful theory by an ugly fact.’’
Harshman et al. 2008:13466
5. Budoucnost a úloha morfologie
Smith a spol. už nevidí velký důvod, proč se nadále zabývat fylogenetickými hypotézami odporujícími monofylii nepštrosích paleognátů. Je pravda, že pro hypotézu monofylie "běžců" sice mluví mnoho analýz morfologických (Mayr & Clarke 2003; Livezey & Zusi 2007; Worthy & Scofield 2012) i molekulárních (Sibley & Ahlquist 1990; García-Moreno & Mindell 2000; Haddrath & Baker 2001) dat, a dokonce ani protikladná evidence ze strany mitochondriální DNA (Phillips et al. 2010) nemusí být průkazná: autoři připomínají, že 37 genů v mitochondrii díky fyzickému propojení reprezentuje jediný "superlokus", takže případný konflikt mezi genovým a druhovým stromem by zůstal neodhalen, a že fylogenetické analýzy mtDNA jsou velice citlivé na taxon sampling a volbu substitučního modelu. Teprve fakt, že Smith et al. (2012) dokázali se zcela nezávislým datasetem dorazit k úplně stejným výsledkům jako Harshman et al. (2008), dává polyfylii "běžců" a monofylii nepštrosích paleognátů náležitou váhu. Ukazuje, že tyto výsledky odrážejí signál rozložený po celém jaderném genomu – ne jenom jedinečné vlastnosti konkrétního datasetu – a nasazení sofistikovaných metod vylučuje možnost, že by za nimi stál jakýkoli známý zdroj nehistorických signálů.
Ponechává tento úspěch molekul prostor pro morfologii? Autoři proto navrhují, aby se budoucí morfologické studie podrobněji zaměřily na rozdíly v ontogenezi jednotlivých "běžců", které by mohly objasnit, jakými mechanizmy tito ptáci dorazili k tak podobnému tělnímu plánu. Stejně tak adaptace na rychlý pohyb po zemi, které daly "běžcům" jejich české jméno, zřejmě vznikly několikrát nezávisle na sobě modifikací tělního plánu silného letce podobného fosilním litornitidům – na tento proces by mohly vrhnout světlo biomechanické studie. Ani slovo ovšem nezaznělo o tom, že by morfologická data mohla pomoct v dalším výzkumu paleognátní fylogeneze. Je zcela očividné, že představují jediný způsob, jak se můžeme něco dozvědět o pozici paleognátních fosilií typu paleotida, diogenornise, eleuterornise, remiornise a četných "litornitidů", kteří jsou stále intenzivněji podezíráni z para- či poly- fylie (Houde 1988; Mayr 2009; Phillips et al. 2010). Postavení těchto taxonů je přitom zajímavé i pro neontology: může pomoct s datací jednotlivých divergencí.
Paleontolog Gerad Mayr ve své skvělé revizi ptačí fylogeneze (Mayr 2011) uznal, že molekuly pomohly v mnoha oblastech, kde morfologie selhala, a že velké datasety sekvenčních dat vykazují daleko příznivější poměr signálu k šumu než srovnatelně rozsáhlé datasety morfologické. Zároveň však učinil i kontroverzní argument, podle nějž se čas od času situace obrací a je to anatomie, která pomáhá rozřešit příbuzenské vztahy i tam, kde molekulára selhala. Jako hlavní příklad takového fenoménu nabídl příbuzenské vztahy uvnitř kladu Strisores ("lelci" včetně svišťounů), což při pohledu zpět nebyla úplně šťastná volba: Mayr zde hodně věřil své dřívější morfologické analýze (Mayr 2010), jejíž výsledky ale zanedlouho po publikaci jeho revizní studie překopali Nesbitt et al. (2011) – je ironické, že své výsledky shledali právě tak robustními jako Mayr (2010) ty svoje. Nešťastně zvolený příklad ale nevyvrací pointu, a je zde lákavá možnost, že morfologie by mohla molekulám pomoci i v jediném zbývajícím otazníku mezi příbuzenskými vztahy žijících paleognátů, totiž s postavením tinam. Je až překvapivé, kolik lze najít znaků spojujících tinamy s nanduy a separujících výslednou skupinu od všech ostatních (žijících) paleognátů, podíváme-li se do literatury dostatečně staré a/nebo obskurní. Ideálním místem, kde začít s hledáním, jsou tak Sibley & Ahlquist (1972), jejichž třísetstránková revize zmiňuje prakticky všechny fylogenetické hypotézy od roku 1800.
Nejsilnějším důkazem pro to, že "běžci" jsou polyfyletickým seskupením v rámci monofyletických paleognátů, je silná molekulární podpora pro skupinu tvořenou všemi paleognáty mimo pštrosů; morfologové se však ke stejné hypotéze nejvíce přiblížili úvahami o podobnostech nanduů a tinam. Beddard (1910) je zaznamenal v trávicím traktu: variace v rámci "Struthiones" podle něj byla vyšší než u mnoha jiných skupin ptáků, ale "possible likeness between Crypturus and Rhea" zmiňuje jako jeden z "certain classificatory results [that] seem to follow from a comparison of the differences exhibited by the intestinal tract" (Beddard 1910:89). Je pravda, že Sibley & Ahlquist (1972) právě na základě Beddardovy klasifikace usoudili, že taxonomická hodnota střevních kliček je prakticky nulová, ale nejde o jediný znak mířící tímto směrem. McDowell (1948) je citován především pro svůj poznatek, že paleognátní stavbu kostěného patra – od dob Huxleyho nejoblíbenější a nejpřesvědčivější znak sjednocující "běžce" navzájem a s tinamami – není možné uspokojivě definovat. Zastánci polyfylie "běžců" a paleognátů typu Mayra nebo Wetmore'a se tohoto zjištění sice rychle chopili, jak ale poznamenávají Sibley & Ahlquist (1972), téměř nikdy neuvedli, že McDowell v rámci paleognátů rozeznal několik skupin, které považoval za monofyletické. Výše citovaným seskupením sloních ptáků a kiviů si nebyl jist, našel však zarážející počet podobností nanduů s tinamami. U obou skupin spolu obě kosti předradličné (prevomery) nedokonale srůstají do velkého nepárového elementu s výraznou "vidlicí" na přední i zadní straně; zadní část horního okraje kosti patrové zespodu vybíhá kousek pod boční okraj zadního konce předradličné kosti a srůstá s ním. Tinamy i nanduové dále sdílí uspořádání patra, v němž kost křídlová (pterygoid) seshora nasedá na prevomer a následně se stáčí dozadu a ven, kde se připojuje na kost patrovou. To je odlišuje od pštrosů (kde křídlová kost nekontaktuje ani prevomer, ani patrovou kost), kasuárovitých (křídlová kost kontaktuje pouze prevomer) i kiviů a sloních ptáků (kde pterygoid kontaktuje předradličnou i patrovou kost najednou, nikoli postupně). Autor dokonce uvádí, že:

Verheyen skutečnost, že "[l]es nandous (Rheiformes) sont apparentés aux tinamous", dokonce konstatoval přímo v titulku jedné ze svých četných studií (Verheyen 1960a). V ní shrnuje morfologické znaky nasvědčující tomu, že nanduové mají blíž k tinamám než k africkým pštrosům (McKinney 1961). Příliš ovšem nepomáhá, že ještě tentýž rok publikoval studii, která naopak argumentovala pro zařazení tinam do hrabavých (Verheyen 1960b).
Parkes & Clark (1966) věřili, že tinamy reprezentují morfologicky nejméně odvozené zástupce paleognátů (což se vzhledem k jejich schopnosti letu zdá být rozumné), a navrhli, že jednotlivé linie "běžců" se vyvinuly z "čeledi" jakýchsi proto-tinam několika nezávislými ztrátami letu. Takový scénář perfektně odpovídá tomu, co navrhují Harshman et al. (2008) a Smith et al. (2012); jediný rozdíl je v tom, že Parkes a Clark neměli explicitní fylogenetickou hypotézu, kterou by ho podložili. Opět však měli co říct k nanduům a tinamám:
Smith a spol. už nevidí velký důvod, proč se nadále zabývat fylogenetickými hypotézami odporujícími monofylii nepštrosích paleognátů. Je pravda, že pro hypotézu monofylie "běžců" sice mluví mnoho analýz morfologických (Mayr & Clarke 2003; Livezey & Zusi 2007; Worthy & Scofield 2012) i molekulárních (Sibley & Ahlquist 1990; García-Moreno & Mindell 2000; Haddrath & Baker 2001) dat, a dokonce ani protikladná evidence ze strany mitochondriální DNA (Phillips et al. 2010) nemusí být průkazná: autoři připomínají, že 37 genů v mitochondrii díky fyzickému propojení reprezentuje jediný "superlokus", takže případný konflikt mezi genovým a druhovým stromem by zůstal neodhalen, a že fylogenetické analýzy mtDNA jsou velice citlivé na taxon sampling a volbu substitučního modelu. Teprve fakt, že Smith et al. (2012) dokázali se zcela nezávislým datasetem dorazit k úplně stejným výsledkům jako Harshman et al. (2008), dává polyfylii "běžců" a monofylii nepštrosích paleognátů náležitou váhu. Ukazuje, že tyto výsledky odrážejí signál rozložený po celém jaderném genomu – ne jenom jedinečné vlastnosti konkrétního datasetu – a nasazení sofistikovaných metod vylučuje možnost, že by za nimi stál jakýkoli známý zdroj nehistorických signálů.
Ponechává tento úspěch molekul prostor pro morfologii? Autoři proto navrhují, aby se budoucí morfologické studie podrobněji zaměřily na rozdíly v ontogenezi jednotlivých "běžců", které by mohly objasnit, jakými mechanizmy tito ptáci dorazili k tak podobnému tělnímu plánu. Stejně tak adaptace na rychlý pohyb po zemi, které daly "běžcům" jejich české jméno, zřejmě vznikly několikrát nezávisle na sobě modifikací tělního plánu silného letce podobného fosilním litornitidům – na tento proces by mohly vrhnout světlo biomechanické studie. Ani slovo ovšem nezaznělo o tom, že by morfologická data mohla pomoct v dalším výzkumu paleognátní fylogeneze. Je zcela očividné, že představují jediný způsob, jak se můžeme něco dozvědět o pozici paleognátních fosilií typu paleotida, diogenornise, eleuterornise, remiornise a četných "litornitidů", kteří jsou stále intenzivněji podezíráni z para- či poly- fylie (Houde 1988; Mayr 2009; Phillips et al. 2010). Postavení těchto taxonů je přitom zajímavé i pro neontology: může pomoct s datací jednotlivých divergencí.
Paleontolog Gerad Mayr ve své skvělé revizi ptačí fylogeneze (Mayr 2011) uznal, že molekuly pomohly v mnoha oblastech, kde morfologie selhala, a že velké datasety sekvenčních dat vykazují daleko příznivější poměr signálu k šumu než srovnatelně rozsáhlé datasety morfologické. Zároveň však učinil i kontroverzní argument, podle nějž se čas od času situace obrací a je to anatomie, která pomáhá rozřešit příbuzenské vztahy i tam, kde molekulára selhala. Jako hlavní příklad takového fenoménu nabídl příbuzenské vztahy uvnitř kladu Strisores ("lelci" včetně svišťounů), což při pohledu zpět nebyla úplně šťastná volba: Mayr zde hodně věřil své dřívější morfologické analýze (Mayr 2010), jejíž výsledky ale zanedlouho po publikaci jeho revizní studie překopali Nesbitt et al. (2011) – je ironické, že své výsledky shledali právě tak robustními jako Mayr (2010) ty svoje. Nešťastně zvolený příklad ale nevyvrací pointu, a je zde lákavá možnost, že morfologie by mohla molekulám pomoci i v jediném zbývajícím otazníku mezi příbuzenskými vztahy žijících paleognátů, totiž s postavením tinam. Je až překvapivé, kolik lze najít znaků spojujících tinamy s nanduy a separujících výslednou skupinu od všech ostatních (žijících) paleognátů, podíváme-li se do literatury dostatečně staré a/nebo obskurní. Ideálním místem, kde začít s hledáním, jsou tak Sibley & Ahlquist (1972), jejichž třísetstránková revize zmiňuje prakticky všechny fylogenetické hypotézy od roku 1800.
Nejsilnějším důkazem pro to, že "běžci" jsou polyfyletickým seskupením v rámci monofyletických paleognátů, je silná molekulární podpora pro skupinu tvořenou všemi paleognáty mimo pštrosů; morfologové se však ke stejné hypotéze nejvíce přiblížili úvahami o podobnostech nanduů a tinam. Beddard (1910) je zaznamenal v trávicím traktu: variace v rámci "Struthiones" podle něj byla vyšší než u mnoha jiných skupin ptáků, ale "possible likeness between Crypturus and Rhea" zmiňuje jako jeden z "certain classificatory results [that] seem to follow from a comparison of the differences exhibited by the intestinal tract" (Beddard 1910:89). Je pravda, že Sibley & Ahlquist (1972) právě na základě Beddardovy klasifikace usoudili, že taxonomická hodnota střevních kliček je prakticky nulová, ale nejde o jediný znak mířící tímto směrem. McDowell (1948) je citován především pro svůj poznatek, že paleognátní stavbu kostěného patra – od dob Huxleyho nejoblíbenější a nejpřesvědčivější znak sjednocující "běžce" navzájem a s tinamami – není možné uspokojivě definovat. Zastánci polyfylie "běžců" a paleognátů typu Mayra nebo Wetmore'a se tohoto zjištění sice rychle chopili, jak ale poznamenávají Sibley & Ahlquist (1972), téměř nikdy neuvedli, že McDowell v rámci paleognátů rozeznal několik skupin, které považoval za monofyletické. Výše citovaným seskupením sloních ptáků a kiviů si nebyl jist, našel však zarážející počet podobností nanduů s tinamami. U obou skupin spolu obě kosti předradličné (prevomery) nedokonale srůstají do velkého nepárového elementu s výraznou "vidlicí" na přední i zadní straně; zadní část horního okraje kosti patrové zespodu vybíhá kousek pod boční okraj zadního konce předradličné kosti a srůstá s ním. Tinamy i nanduové dále sdílí uspořádání patra, v němž kost křídlová (pterygoid) seshora nasedá na prevomer a následně se stáčí dozadu a ven, kde se připojuje na kost patrovou. To je odlišuje od pštrosů (kde křídlová kost nekontaktuje ani prevomer, ani patrovou kost), kasuárovitých (křídlová kost kontaktuje pouze prevomer) i kiviů a sloních ptáků (kde pterygoid kontaktuje předradličnou i patrovou kost najednou, nikoli postupně). Autor dokonce uvádí, že:
The similarity of the palate of the Rheidae to that of the Tinamidae is also borne out by similarities in the nasals, lacrymals, calvarium, costal processes, hypotarsus, sculpture of leg-bones, coiling of gut, etc. This resemblance is so great, in fact, that close phylogenetic relationship seems unavoidable.A dále v textu:
McDowell 1948:540
It would also seem wise to simplify the classification of this group by the merging of certain orders which seem, on the basis of palatal pattern, to be closely related. Thus:Je pravda, že korespondence mezi McDowellovými závěry a výsledky molekulárních fylogenetických analýz není úplná. Ptáky moa nezařadil do stejné skupiny jako tinamy a nanduy; místo toho je připojil ke sloním ptákům a kivům pod Apterygiformes. (Stojí ovšem za zmínku, že právě ptáci moa se mu z diagnózy skupiny nejčastěji vymykali.)
Order Tinamiformes to include the Rheidae in addition to the tinamous, thus eliminating the order ‘Rheiformes’ from the list.
McDowell 1948:547

Kostěné patro nandu pampového (Rhea americana, A) a tinamy inambu (Rhynchotus, B) při pohledu seshora (vlevo) a zespoda (vpravo). Oba taxony v této oblasti sdílí řadu znaků, které u jiných paleognátů nenacházíme – zvláštní skloubení kosti křídlové s kostí předradličnou a kostí patrovou, morfologii předradličné kosti a tvar bazipterygoidních a parasfenoidních výběžků. Většina rozdílů je pouze zdánlivá a způsobená tím, že patro tinam je delší a užší. Význam zkratek: (bp) bazipterygoidní výběžek, (mp) patrový výběžek maxilly, (p) kost křídlová, (pa) kost patrová, (pm) předčelistní kost, (pv) prevomer, (ps) parasfenoid, (q) kost čtvercová. (Modifikováno z McDowell 1948: Text-figs. 1 a 2)
Verheyen skutečnost, že "[l]es nandous (Rheiformes) sont apparentés aux tinamous", dokonce konstatoval přímo v titulku jedné ze svých četných studií (Verheyen 1960a). V ní shrnuje morfologické znaky nasvědčující tomu, že nanduové mají blíž k tinamám než k africkým pštrosům (McKinney 1961). Příliš ovšem nepomáhá, že ještě tentýž rok publikoval studii, která naopak argumentovala pro zařazení tinam do hrabavých (Verheyen 1960b).
Parkes & Clark (1966) věřili, že tinamy reprezentují morfologicky nejméně odvozené zástupce paleognátů (což se vzhledem k jejich schopnosti letu zdá být rozumné), a navrhli, že jednotlivé linie "běžců" se vyvinuly z "čeledi" jakýchsi proto-tinam několika nezávislými ztrátami letu. Takový scénář perfektně odpovídá tomu, co navrhují Harshman et al. (2008) a Smith et al. (2012); jediný rozdíl je v tom, že Parkes a Clark neměli explicitní fylogenetickou hypotézu, kterou by ho podložili. Opět však měli co říct k nanduům a tinamám:
The situation in the New World is particularly interesting. It appears reasonable to assume that the rheas, of all of the large flightless birds, have attained the “ratite grade” most recently. Their wings are the least reduced, and they show many resemblances to tinamous, leading McDowell ( 1948) to combine the Rheidae and Tinamidae into a single order. It seems to us to be probable that the rheas evolved flightlessness in South America, possibly from ancestors appreciably nearer the tinamou than the proto-tinamou level. Thus they retain ancestral characteristics common to the whole group (cf. the rhamphothecal structure), but share certain special characteristics with the only other New World group, the tinamous.
Parkes & Clark 1966:468
Jehl (1971) zkoumal barevné vzorce v prachovém peří mladých paleognátů a dospěl k závěru, že tinamy nemají v tomto ohledu s "běžci" prakticky nic společného. Zajímavější je však jeho zmínka o tom, že "[t]he attempt to establish a phylogenetic link between the ratites and tinamous, and particularly between the rheas and tinamous, dates at least to the early 19th century, and the downy young have been used to give some support to that view." (Jehl 1971:300). Sám autor přiznal, že ačkoli jeho data takové příbuzenství nepodporují, nestačí k tomu, aby mohlo být zamítnuto. Klad (nanduové + tinamy) vyčetli z jazykového aparátu i Bock & Bühler (1990), jejichž pětistránkový abstrakt v obskurním sborníku stál za zrodem dnešní podoby hypotézy o "běžčí" polyfylii (viz přechozí díl článku). Tomlinson (2000) mnohá pozorování Bocka a Bühlera opravila a jejich závěry kritizovala – místo toho přišla s vlastní, o nic méně revoluční hypotézou, podle níž monofyletičtí "běžci" tvoří sesterskou skupinu ke kladu složenému z tinam a neognátů. I tak ale ve své práci uvedla četné podobností tinam a jejich jihoamerických příbuzných: tvar jazyka i charakteristiky jazykové sliznice jsou u obou taxonů v podstatně shodné a paraglossale (chrupavčitá součást jazylky, viz minulý článek) některých tinam je dokonce podobnější témuž elementů u nanduů než u jiných tinam. Podobnosti vykazuje i vnější jazylkojazykové svalstvo. Zatahování jazyka je u nanduů i tinam realizováno nepřímým působením svalů na jazylkové rohy, zatímco u emu jiná sada svalů působí přímo na ceratobranchiály a urohyale a u pštrosa přímo na ceratobranchiály a hrtan (Tomlinson 2000:379).
Jelikož nepštrosí paleognáti nejsou tvrdá polytomie, vztahy tinam dříve či později odhalíme – jen budeme muset identifikovat a osekvenovat potřebné desítky (či lépe stovky) nových vhodných lokusů. Výše uvedené studie se ale zdají napovídat, že existuje i jednodušší cesta, totiž detailní fylogenetická analýza lebečních a měkkotkáňových znaků, které ornitologové za poslední století dokázali nashromáždit.
6. Acknowledgments
Děkuji Janu Zrzavému za přístup k fulltextu studie Smithe a kolektivu, Nicolási Diezovi za přístup k řadě dalších prací zmiňovaných v tomto článku, a Alexanderu Suhovi za to, že mě upozornil na abstrakt Bakera a Haddratha.
Zdroje:
Jelikož nepštrosí paleognáti nejsou tvrdá polytomie, vztahy tinam dříve či později odhalíme – jen budeme muset identifikovat a osekvenovat potřebné desítky (či lépe stovky) nových vhodných lokusů. Výše uvedené studie se ale zdají napovídat, že existuje i jednodušší cesta, totiž detailní fylogenetická analýza lebečních a měkkotkáňových znaků, které ornitologové za poslední století dokázali nashromáždit.
6. Acknowledgments
Děkuji Janu Zrzavému za přístup k fulltextu studie Smithe a kolektivu, Nicolási Diezovi za přístup k řadě dalších prací zmiňovaných v tomto článku, a Alexanderu Suhovi za to, že mě upozornil na abstrakt Bakera a Haddratha.
Zdroje:
- http://www.stat.wisc.edu/~ane/bucky/prior.html
- http://www.dnabased.com/Bioinformatika/Prednasky/.../Bayes_web_soubory/frame.htm
- Allentoft ME, Rawlence NJ 2011 Moa’s Ark or volant ghosts of Gondwana? Insights from nineteen years of ancient DNA research on the extinct moa (Aves: Dinornithiformes) of New Zealand. Ann Anat 194: 36–51
- Alvarenga H 2010 Diogenornis fragilis Alvarenga, 1985, restudied: a South American ratite closely related to Casuariidae. 146 in Miyaki CY, Höfling E, Donatelli RJ, eds. Abstracts XXV Int Orn Congr (Campos de Jordão 2010): recurso eletrônico. Campos do Jordão: IOC
- Baker AJ, Haddrath O 2010 Rapid radiations and incomplete lineage sorting tangle the branches of the Avian Tree of Life. 129 in Miyaki CY, Höfling E, Donatelli RJ, eds. Abstracts XXV Int Orn Congr (Campos de Jordão 2010): recurso eletrônico. Campos do Jordão: IOC
- Beddard FE 1910 On the alimentary tract of certain birds and on the mesenteric relations of the intestinal loops. Proc Zool Soc Lond 1910: 47–93
- Bledsoe AH 1988 A phylogenetic analysis of postcranial skeletal characters of the ratite birds. Ann Carnegie Mus 57: 73–90
- Bock WJ, Bühler P 1990 The evolution and biogeographical history of the palaeognathous birds. 31–6 in van den Elzen R, Schuchmann K-L, Schmidt-Koenig K, eds. Current Topics in Avian Biology: Proceedings of the International Centennial Meeting of the Deutsche Ornithologen-Gesellschaft (Bonn 1988). Bonn: Verlag der Deutschen Ornithologen-Gesellschaft
- Bunce M, Worthy TH, Phillips MJ, Holdaway RN, Willerslev E, Haile J, Shapiro B, Scofield RP, Drummond A, Kamp PJJ, Cooper A 2009 The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography. Proc Natl Acad Sci USA 106(49): 20646–51
- Chojnowski JL, Kimball RT, Braun EL 2008 Introns outperform exons in analyses of basal avian phylogeny using clathrin heavy chain genes. Gene 410(1): 89–96
- Cooper A, Lalueza-Fox C, Anderson S, Rambaut A, Austin J, Ward R 2001 Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution. Nature 409(6821): 704–7
- Dawkins R 2004[2008] [Příběh předka: pouť k úsvitu života.] Praha: Academia [Transl. by Gabajová Z]
- Degnan JH, Rosenberg NA 2006. Discordance of species trees with their most likely gene trees. PLoS Genetics 2(5): e68
- Faircloth BC, McCormack JE, Crawford NG, Harvey MG, Brumfield RT, Glenn TC 2012 Ultraconserved elements anchor thousands of genetic markers spanning multiple evolutionary timescales. Syst Biol 61(5): 717–26
- García-Moreno J, Mindell DP 2000 Rooting a phylogeny with homologous genes on opposite sex chromosomes (gametologs): A case study using avian CHD. Mol Biol Evol 17(12): 1826–32
- Hackett SJ, Kimball RT, Reddy S, Bowie RC, Braun EL, Braun MJ, Chojnowski JL, Cox WA, Han K, Harshman J, Huddleston CJ, Marks BD, Miglia KJ, Moore WS, Sheldon FH, Steadman DW, Witt CC, Yuri T 2008 A phylogenomic study of birds reveals their evolutionary history. Science 320(5884): 1763–8
- Haddrath O, Baker AJ 2001 Complete mitochondrial DNA genome sequences of extinct birds: Ratite phylogenetics and the vicariance biogeography hypothesis. Proc R Soc Lond B 268(1470): 939–45
- Harshman J, Braun EL, Braun MJ, Huddleston CJ, Bowie RCK, Chojnwoski JL, Hackett SJ, Han K-L, Kimball RT, Marks BD, Miglia KJ, Moore WS, Reddy S, Sheldon FH, Steadman DW, Steppan SJ, Witt CC, Yuri T 2008 Phylogenomic evidence for multiple losses of flight in ratite birds. Proc Natl Acad Sci USA 105(36): 13462–7
- Hope S 2002 The Mesozoic radiation of Neornithes. 339–88 in Chiappe LM, Witmer LM, eds. Mesozoic birds: Above the Heads of Dinosaurs. Berkeley, CA: Univ of California Press
- Houde P 1988 Palaeognathous birds from the early Tertiary of the Northern Hemisphere. Publ Nuttall Ornithol Club 22: i–vii, 1–148
- Houde P, Haubold H 1987 Palaeotis weigelti restudies: a small Eocene ostrich (Aves: Struthionifomes). Palaeovertebr 17: 27–42
- Huelsenbeck JP, Rannala B 2004 Frequentist properties of Bayesian posterior probabilities of phylogenetic trees under simple and complex substitution models. Syst Biol 53(6): 904–13
- Jehl JR Jr 1971 The color patterns of downy young ratites and tinamous. Trans S Diego Soc Nat Hist 16: 292–301
- Johnston P 2011 New morphological evidence supports congruent phylogenies and Gondwana vicariance for palaeognathous birds. Zool J Linn Soc 163(3): 959–82
- Kolaczkowski B, Thornton JW 2008 A mixed branch length model of heterotachy improves phylogenetic accuracy. Mol Biol Evol 25(6): 1054-66
- Kubatko LS, Degnan JH 2007 Inconsistency of phylogenetic estimates from concatenated data under coalescence. Syst Biol 56(1): 17–24
- Livezey BC, Zusi RL 2007 Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy: II. – Analysis and discussion. Zool J Linn Soc 149(1): 1–95
- Matsen FA, Steel M 2007 Phylogenetic mixtures on a single tree can mimic a tree of another topology. Syst Biol 56(5): 767–75
- Maxwell EE, Larsson HCE 2009 Comparative ossification sequence and skeletal development of the postcranium of palaeognathous birds (Aves: Palaeognathae). Zool J Linn Soc 157(1): 169–96
- Mayr G 2009 Paleogene Fossil Birds. Heidelberg: Springer
- Mayr G 2010 Phylogenetic relationships of the paraphyletic ‘caprimulgiform’ birds (nightjars and allies). J Zool Syst Evol Res 48(2): 126–37
- Mayr G 2011 Metaves, Mirandornithes, Strisores and other novelties – a critical review of the higher-level phylogeny of neornithine birds. J Zool Syst Evol Res 49(1): 58–76
- Mayr G, Clarke JA 2003 The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19: 527–53
- McDowell S 1948 The bony palate of birds. Part 1. The Palaeognathae. Auk. 65: 520–49
- McKinney F 1961 Recent literature. Auk 78(3): 455–68
- Morgan-Richards M, Trewick SA, Bartosch-Härlid A, Kardailsky O, Phillips MJ, McLenachan PA, Penny D 2008 Bird evolution: testing the Metaves clade with six new mitochondrial genomes. BMC Evol Biol 8: 20
- Nesbitt SJ, Ksepka DT, Clarke JA 2011 Podargiform affinities and the enigmatic Fluvioviridavis platyrhamphus and the early diversification of Strisores (‘‘Caprimulgiformes’’ + Apodiformes). PLoS ONE 6(11): e26350
- Parkes KC, Clark GA 1966 An additional character linking ratites and tinamous and an interpretation of their monophyly. Condor 68(5): 459
- Parris DC, Hope S 2002 New interpretations of birds from the Navesink and Hornerstown formations, New Jersey, USA (Aves: Neornithes). 113–24 in Zhou Z-H, Zhang F-C, eds. Proc 5th Symp Soc Avian Paleont Evol (Beijing 2000). Beijing: Science Press
- Phillips MJ, Gibb GC, Crimp EA, Penny D 2010 Tinamous and moa flock together: mitochondrial genome sequence analysis reveals independent losses of flight among ratites. Syst Biol 59(1): 90–107
- Poe S, Chubb AL 2004 Birds in a bush: five genes indicate explosive evolution of avian orders. Evolution 58(2): 404–15
- Ronquist F, Deans AR 2010 Bayesian phylogenetics and its influence on insect systematics. Annu Rev Entomol 55:189-206
- Sibley CG, Ahlquist JE 1972 A comparative study of egg-white proteins of non-passerine birds. Bull Peabody Mus Nat Hist 39: 1–276
- Sibley CG, Ahlquist JE 1990 Phylogeny and classification of birds: a study in molecular evolution. New Haven: Yale Univ Press
- Smith JV, Braun EL, Kimball RT 2012 Ratite non-monophyly: Independent evidence from 40 novel loci. Syst Biol doi:10.1093/sysbio/sys067
- Steadman DW 1995 Prehistoric extinctions of pacific island birds: biodiversity meets zooarchaeology. Science 267(5201): 1123–31
- Tomlinson CAB 2000 Feeding in paleognathous birds. 359–94 in Schwenk K, ed. Feeding: Form, Function, and Evolution in Tetrapod Vertebrates. San Diego: Acad Press
- Tuffley C, Steel M 1998 Modelling the covarion hypothesis of nucleotide substitution. Math Biosci 147: 63–91
- Verheyen R 1960a Les nandous (Rheiformes) sont apparentés aux tinamous (Tinamidae/Galliformes). Le Gerfaut 50: 289–93
- Verheyen R 1960b Les tinamous dans les systèmes ornithologiques. Bull Inst Roy Sci Nat Belgique 36(1): 1–11
- Wang N, Braun EL, Kimball RT 2011 Testing hypotheses about the sister group of the Passeriformes using an independent 30-locus data set. Mol Biol Evol 29(2): 737–50
- Whiting MF, Bradler S, Maxwell T 2003 Loss and recovery of wings in stick insects. Nature 421(6920): 264–7
- Worthy TH, Scofield RP 2012 Twenty-first century advances in knowledge of the biology of moa (Aves: Dinornithiformes): a new morphological analysis and moa diagnoses revised. New Zealand J Zool 39(2): 87–153



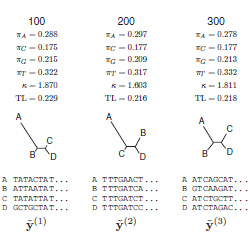








0 komentářů:
Okomentovat
Povolené tagy:
- <b>tučně</b> = tučně
- <i>kurzíva</i> = kurzíva
- <a href="http://pan-aves.blogspot.com/">pan-Aves</a> = pan-Aves
Podporuje $\mathrm{\LaTeX}$ pro matematické vzorce.